- Wissen
25 Jahre Wachstumsentwicklung in 19 Schweizer Wäldern unter zunehmender Trockenheit
26.02.2025
Abstract
Wälder sind eine wichtige Kohlenstoffsenke, stehen jedoch durch sich häufende Klimaextreme wie Hitze und Trockenheit zunehmend unter Druck. Nach 25 Jahren Monitoring an den 19 Standorten des Programms «Langfristige Waldökosystem-Forschung» zeigen sich Veränderungen in der Waldstruktur und im Wachstum. Die Wälder wurden generell dichter und älter. Das Wachstum nahm sowohl in den Tief- als auch in den Hochlagen ab, vor allem bei Fichte, Tanne und Buche. Das Wachstum korreliert auf Einzelbaum- und auf Bestandesebene mit Bestandesstruktur (Bestandesdichte und Alter) und negativ mit Temperatur und Trockenheit. Verglichen mit den 1990er-Jahren weisen die Wälder an den untersuchten Standorten heute eine geringere Produktivität bei gleicher Bestandesdichte auf, was auf eine Abnahme der Standortskapazität hinweist.
Keywords: forest growth, stand density, net primary productivity, carbon uptake, drought, dendrometer
Schweiz Z Forstwesen 176 (2): 77–83. https://doi.org/10.3188/szf.2025.0077
* Zürcherstrasse 111, CH-8903 Birmensdorf, E-Mail sophia.etzold@wsl.ch
Bäume nehmen Kohlenstoff in Form von CO2 aus der Atmosphäre auf, wandeln es mithilfe des Sonnenlichts in Zucker um und bauen unter anderem damit Holz auf. Ein Kubikmeter Holz speichert je nach Baumart etwa 0.2–0.4 Tonnen Kohlenstoff, was etwa einer Tonne aufgenommenem CO2 entspricht. Wälder entziehen der Atmosphäre also grosse Mengen an CO2. Darüber hinaus tragen sie zur Abkühlung des Klimas bei und bieten Schutz vor Naturgefahren. Gleichzeitig stehen sie zunehmend unter Druck wegen Trockenheit, Hitze und des gehäuften Auftretens von Extremereignissen wie Stürme, Waldbrände und Überschwemmungen. Hinzu kommt die Verbreitung von Schädlingen wie des Borkenkäfers. Neueste Berichte über Waldsterben durch Trockenheit in Europa führten zur Erkenntnis, dass neue innovative Ansätze in der Waldbewirtschaftung und im Naturschutz notwendig sind (IPCC 2022). Langfristige Monitoringdaten sind eine unentbehrliche Grundlage, um die Auswirkungen des Klimawandels auf den Waldzustand zu bewerten und Anpassungsstrategien zu entwickeln.
Die Beziehung zwischen dem Waldwachstum bzw. der Waldgesundheit und abiotischen Einflussfaktoren ist oftmals nicht linear und zudem standortabhängig. Während in den Tieflagen erhöhte Temperaturen und Trockenheit das Wachstum bremsen, wird in den höheren Lagen eine Wachstumszunahme durch höhere Temperaturen und eine verlängerte Vegetationsperiode erwartet (Jolly et al 2005, Dulamsuren et al 2017). Neben dem Klima spielt auch die Nährstoffversorgung eine wichtige Rolle. Erhöhte Stickstoffeinträge können zunächst wachstumsfördernd wirken, aber bei Überversorgung negative Effekte auf die Waldgesundheit haben (Etzold et al 2020).
Seit 1995 wird das Wachstum der Bäume auf den Flächen des Programms «Langfristige Waldökosystem-Forschung» (LWF) in regelmässigen Abständen von etwa fünf Jahren gemessen. Inzwischen liegen Zeitreihen über 20–25 Jahre vor. 25 Jahre sind in einem Baumleben zwar nur ein kurzer Zeitraum, dennoch waren die letzten Jahrzehnte durch zahlreiche Extremereignisse und zunehmende Trockenheit (NCCS 2018) geprägt. In diesem Artikel untersuchen wir die Entwicklung der Produktivität auf den 19 LWF-Flächen während der letzten Dekaden. Wir erwarten, dass Wälder auf trockenen und warmen Standorten Produktivitätsverluste zeigen, während Wälder in höheren Lagen von wärmeren Temperaturen und längeren Wachstumsperioden profitieren und ihr Wachstum steigern können. Wir prüfen, welche Einflussfaktoren (z.B. Bestandesstruktur [Dichte und Alter], Klima und Nährstoffe) das Wachstum der Hauptbaumarten und die Produktivität der Bestände am besten erklären.
Monitoring des Baumwachstums auf den LWF-Flächen
Die LWF-Flächen wurden in den 1990er-Jahren angelegt. Das Ziel war, Veränderungen des Waldzustandes frühzeitig zu erkennen und externe Einflüsse wie Klima, Nähr- und Schadstoffeinträge und ihre Auswirkung auf das Ökosystem Wald zu erfassen (LWF 2024). Das Wachstum ist hierbei ein wichtiger Indikator für den Waldzustand und die Waldgesundheit (Dobbertin 2005).
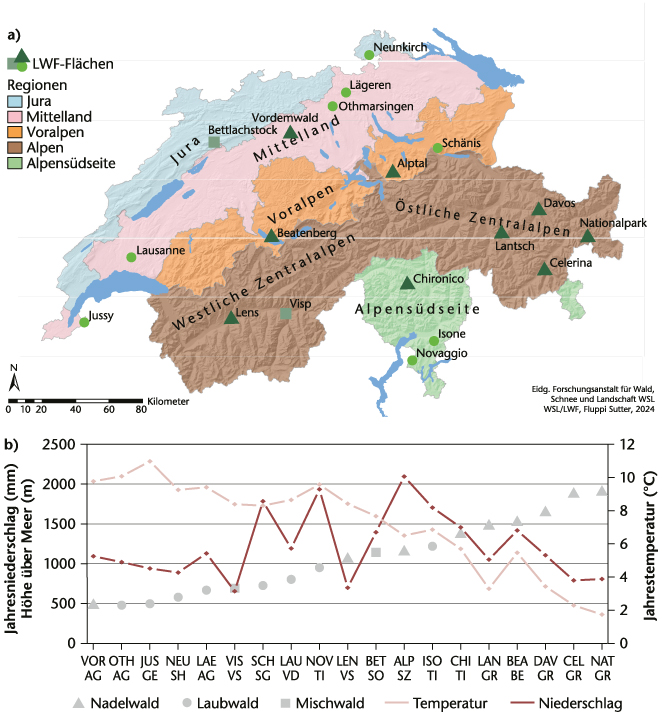
Die 19 LWF-Flächen wurden dahingehend ausgewählt, dass sie charakteristische Waldökosysteme der Schweiz abdecken. Die Entwicklungen auf den LWF-Flächen sind jedoch nicht unbedingt repräsentativ für die ganze Schweiz. Insbesondere nicht für diejenigen Baumarten, die nur auf einzelnen oder wenigen LWF-Flächen vorkommen, da es grosse standörtliche Unterschiede gibt. Daher versuchten wir, unsere Ergebnisse den Entwicklungen des Landesforstinventars (LFI 2023) gegenüberzustellen, dessen Daten in einem systematischen 1.4-×-1.4-km-Netz erhoben werden. Zu den LWF-Wäldern gehören neun Nadelwälder, acht Laubwälder und zwei Mischwälder, die entlang eines Höhengradienten von 500 bis 1900 m ü.M. liegen und somit sehr unterschiedlichen Klimabedingungen ausgesetzt sind (Abbildung 1). Es gibt warme trockene Standorte, vor allem im Wallis, Jura und Mittelland, feuchte Standorte im Tessin und an der Alpennordflanke sowie kühle, aber trockene Standorte in höheren Lagen. Die Wälder unterscheiden sich stark hinsichtlich Bestandesdichte, Alter und Artenzusammensetzung (vgl. Tabelle 1 in Etzold et al 2014, Tabelle 1 in Thimonier et al 2025).
Das Wachstum der Bäume wird etwa alle fünf Jahre in Inventuren erfasst. Dabei werden die Brusthöhendurchmesser der Stämme auf 1.3 m über Boden (BHD) und die Höhe aller Bäume auf einer definierten Fläche gemessen. Begleitend dazu wird die Vitalität (lebend/tot) der Bäume erfasst. Die LWF-Flächen bestehen meistens aus zwei Teilflächen (A und B, insgesamt ca. 1 ha gross) und zwei Subflächen (je ca. 0.25 ha) innerhalb der Teilflächen. Auf den Teilflächen werden alle Bäume mit BHD >12 cm aufgenommen, auf den Subflächen auch solche mit BHD >5 cm.
Ausgehend von BHD und Baumhöhe wurden die verschiedenen Baumkompartimente (Schaft- und Derbholz, Wurzeln, Feinwurzeln, Äste und Zweige sowie Blätter und Nadeln) mit allometrischen Funktionen (Kaufmann 2001) geschätzt und, falls vorhanden, mit gemessenen Werten (Blätter und Nadeln) validiert. Der Holzvorrat eines Waldes ist die Summe des Schaftholzes (in m3/ha) aller lebenden Bäume. Die Produktivität des Waldes (Biomasse, in kg/ha/Jahr) ergibt sich aus der Summe des ober- und unterirdischen Baumwachstums zwischen zwei Inventuren (Abgänge werden hier nicht berücksichtigt). Biomasse (BM) wurde in Kohlenstoffaufnahme (Nettoprimärproduktion [NPP] in g C/m2/Jahr) umgerechnet, mit der Annahme, dass 1 kg BM 0.5 kg Kohlenstoff entspricht. Zusätzlich dazu wurde der Basalflächenzuwachs (BFZ in cm2/ha) erfasst. Alle allometrischen Funktionen sind in Etzold et al (2014) detailliert beschrieben. Insgesamt wurden während des gesamten Zeitraums 21 219 Baumindividuen erfasst, von denen ca. 3780 während der Monitoringzeit starben oder gefällt wurden und ca. 3300 neu einwuchsen.
Seit dem Jahr 2000 wurde auf jeder LWF-Fläche der jährliche Stammradiuszuwachs von jeweils etwa zehn Bäumen der dominierenden Baumarten mit permanenten manuellen Umfangmessbändern (Dendrometer D1, Meter Group, München) erfasst. Anhand des hergeleiteten Basalflächenanteils der jeweiligen Baumart innerhalb der LWF-Fläche und ihres relativen jährlichen Stammradiuszuwachses konnten wir die fünfjährige Produktivität des Waldes auf eine jährliche Basis skalieren.
Für alle LWF-Flächen und Inventurperioden berechneten wir den Bestandesdichteindex nach Reinecke (1933, Stand Density Index [SDI]) und verwendeten den mittleren BHD als Schätzung für das Bestandesalter. Temperatur, Niederschlag, Bodenwasser (Matrixpotenzial in kPa) und ein Trockenstressindikator (das Verhältnis von aktueller zu potenzieller Transpiration) wurden für alle Flächen modelliert (Meteodaten: Meteotest 2020, Bodenwasser: Meusburger et al 2025). Diese Modelle wurden in saisonale Perioden (Frühling: März–Mai, MAM, Sommer: Juni–August, JJA) oder über die Dauer der Vegetationsperiode (April–Oktober) sowie über die Dauer der jeweiligen Inventurperioden aggregiert. Der Stickstoffeintrag über trockene und nasse Deposition wurde in Fünfjahresschritten von 2000 bis 2020 für alle LWF-Flächen modelliert (Rihm & Achermann 2016). Alle zwei Jahre wurden Stickstoff- und Phosphorgehalte in Blättern und Nadeln auf den LWF-Flächen bestimmt (Meusburger et al 2025). Für Nadelbäume wird hier nur der Nährstoffgehalt in diesjährigen Nadeln gezeigt.
Produktivität auf den LWF-Standorten
Fünfjahresinventuren
In den letzten 25 Jahren nahmen sowohl der Holzvorrat als auch die Bestandesdichte und der mittlere BHD auf den meisten LWF-Flächen zu (p <0.01), ausser im Nationalpark (NAT) und in Visp (VIS) (Abbildung 2). Der Holzzuwachs durch Wachstum übertraf die Verluste durch Abgänge und forstliche Nutzung. Die generelle Zunahme des Holzvorrats und der Bestandesdichte ist auf die selektive und geringe Bewirtschaftung der LWF-Flächen zurückzuführen. Allerdings schwächte sich die Zunahme des Holzvorrats auf neun Flächen im Jura, im Mittelland und in den Voralpen im Vergleich zu früheren Jahren deutlich ab. Auch die Daten der fünften Erhebung des Landesforstinventars (LFI) von 2018 bis 2022 und der vierten Bundeswaldinventur in Deutschland im Jahr 2022 zeigen, dass der Holzvorrat in der Schweiz vor allem im Jura und im Mittelland stark abnahm (LFI 2023), in Deutschland vor allem in Nadelwäldern (BMEL 2024).
![Abb 2 Kohlenstoffaufnahme durch Wachstum (NPP in g C/m2/Jahr, abgesetzt [hellrot] jeweils der Anteil der jungen Bäume von 5 bis 12 cm, auf den Subflächen aufgenommen) sowie Holzvorrat (m3/ha/Jahr), Bestandesdichteindex (SDI) und mittlerer BHD (cm) pro LWF-Fläche über die letzten 20–25 Jahre. Die farbigen Balken unter den Namen geben die Region an (vgl. Abbildung 3).](/fileadmin/article_images/159595.svg)
Die LWF-Wälder nahmen durch Wachstum über die letzten 25 Jahre im Durchschnitt 450 g C/m2/Jahr bei einem Basalflächenwachstum von 447 cm2/m2/Jahr auf. Im Mittelland wurde die höchste Produktivität (674 g C/m2/Jahr) gemessen, in den Alpen die geringste (270 g C /m2/Jahr) (Abbildung 2), was mit den LFI-Daten übereinstimmt. Zwischen 2004 und 2016 (Inventurperioden III und IV) war die Produktivität am höchsten (p <0.05), nahm nach 2014 aber auf den meisten Flächen ab und lag unter dem jeweiligen Durchschnitt.
Der Anteil der Mortalität und die Verjüngung wurden als prozentualer Anteil zum Gesamtvolumen berechnet (Abbildung 3). Die beiden Flächen NAT und VIS wiesen eine Mortalität von über 40% des Gesamtvolumens auf (im Durchschnitt über alle Inventurperioden), bedingt durch Pilzbefall (NAT, Dobbertin et al 2001) und extreme Trockenheit (VIS, Rigling et al 2013, Hunziker et al 2022). Bei allen anderen Flächen lag die (konkurrenzbedingte) Mortalität unter 10% des Gesamtvolumens. Bei den meisten Flächen war die Verjüngung (Bäume mit BHD zwischen 5 und 12 cm) prozentual ähnlich hoch wie die Mortalität. Zudem hatte die Verjüngung auf Flächen mit hoher Sterblichkeit einen grösseren Anteil an der Gesamtzuwachsleistung des Waldes (= NPP aller Bäume <12 cm im Verhältnis zu allen Bäumen auf den Subflächen). Bei elf der Flächen lag er unter 10%. Den höchsten Anteil am Gesamtzuwachs hatte die Verjüngung in NAT (53%) und in VIS (81%).

Jährliches Wachstum
Über alle Standorte ist eine Abnahme des Bodenwasserpotenzials seit dem Jahr 2000 zu sehen (Abbildung 4). Die jährliche NPP und der BFZ zeigen je nach Baumart und Standort unterschiedliche zeitliche Verläufe. Auf den von Buchen dominierten Standorten im Jura und im Mittelland sowie in den Voralpen ist ein signifikanter Abwärtstrend von –2.4% NPP und –3.7% BFZ pro Jahr zu sehen. Vor allem ab dem Jahr 2014 liegen die meisten Werte unter dem Durchschnitt. Starkes Wachstum trat in den Jahren 2010–2012 auf, was das hohe Wachstum noch während der vierten Inventurperiode von 2009 bis 2016 (vgl. Abbildung 2) erklärt. Auch die fichtendominierten Wälder zeigen einen signifikant abnehmenden Trend, der vor allem ab dem Jahr 2015 sichtbar wird (Abbildung 4). Für die Eichen und Föhrenarten ist kein eindeutiger Trend zu erkennen. Auffällig bei den Föhren sind jedoch die starken jährlichen Schwankungen, die wahrscheinlich auch durch das generell geringe Wachstum zu erklären sind.
Teilt man die Standorte in Hoch- und Tieflagenstandorte (Grenzwert 1000 m ü.M., alle Baumarten berücksichtigt) auf, sehen wir nach 2010 eine signifikante Abnahme des NPPs von 0.018% pro Jahr in den Tieflagen und 0.020% pro Jahr in den Hochlagen (p <0.001). Das entspricht einer mittleren Differenz von –122 bzw. –116 g C/m2/Jahr in diesem Zeitraum. Die Produktivitätsabnahme in den Tieflagen kann auf die trockenen und heissen Sommer zurückgeführt werden. In den Hochlagen erwarteten wir allerdings eine Produktionssteigerung durch wärmere Temperaturen und verlängerte Vegetationsperioden, wie zum Beispiel der Hitzesommer 2003 gezeigt hatte (Jolly et al 2005). Die Abnahme der Produktivität in den Hochlagen liegt jedoch in einem ähnlichen Bereich wie in den Tieflagen oder ist sogar noch ausgeprägter, wenn man berücksichtigt, dass die absolute Produktivität in höheren Lagen generell geringer ist.

Einflussfaktoren auf das Baum- und Waldwachstum
Um die Gründe für das abnehmende Wachstum zu untersuchen, haben wir den fünfjährigen BFZ der Einzelbäume und des gesamten Bestandes mithilfe gemischter Regressionsmodelle analysiert (Abbildung 5). Als erklärende Faktoren wurden Variablen der Bestandesstruktur, des Klimas und der Nährstoffversorgung getestet und für die Einzelbaummodelle zusätzlich der BHD des Baumes, um den Alterseffekt zu berücksichtigen. Dargestellt sind nur jene Variablen, die die engste Beziehung mit dem Wachstum aufwiesen. Bestandesstruktur und Klimabedingungen waren am stärksten mit Wachstum korreliert, während Nährstoffe in Luft und Blättern sowie Bodeneigenschaften (C/N, pH-Wert) nur zum Teil einen signifikanten Zusammenhang zeigten. Mit zunehmender Bestandesdichte nahm das Einzelbaumwachstum durch Konkurrenz um Platz, Licht und andere Ressourcen erwartungsgemäss ab – mit zunehmendem Bestandesalter (mittlerer BHD des Bestandes) nahm es zu. Bei den Klimabedingungen zeigten alle Hauptbaumarten bis auf die Föhre ein ähnliches Bild. Das Wachstum nahm mit zunehmenden Temperaturen ab und stieg mit zunehmender Wasserverfügbarkeit (Niederschlag, Bodenwasser) an. Die Buche reagierte hierbei am stärksten negativ auf Temperatur und positiv auf Bodenwasserverfügbarkeit (repräsentiert über das Matrixpotenzial). Eine hohe Anfälligkeit der Buche auf Trockenheit und Hitze wurde nach dem Jahr 2018 insbesondere auf den flachgründigeren Südhängen des Juras beobachtet (Rohner et al 2021).

Wir interpretieren die Resultate der LWF-Flächen dahingehend, dass die Buche mit zunehmender Trockenheit und Hitze auch auf tiefgründigeren Standorten des Mittellandes an ihre Belastungsgrenze gestossen ist, was sich in den vorliegenden Daten in abnehmendem Wachstum zeigt. Prognosen deuten darauf hin, dass die Buche längerfristig auch im Mittelland an ihre Toleranzgrenze stossen wird und bei zukünftig moderaten Klimaveränderungen ab Mitte des Jahrhunderts keine geeigneten Wuchsbedingungen mehr vorfinden und damit dieses Areal verlieren wird (Trotsiuk et al 2021, Gessler et al 2024). Im Gegensatz dazu korrelierte das Wachstum der Föhrenarten stark positiv mit Temperatur und stark negativ mit Wasserverfügbarkeit. Wir erklären dieses Muster mit der hohen Lage der Föhrenwälder, wie den Flächen NAT und Celerina (CEL) am oberen Ende des Höhengradienten, wo das Wachstum durch tiefe Temperaturen begrenzt wird. Stickstoffdeposition war für Tanne und Eiche positiv mit Wachstum korreliert, für Föhre dagegen negativ. Das ist insofern erstaunlich, da die Stickstoff-Blattspiegelwerte der Föhren auf den LWF-Flächen im Mangelbereich liegen (vgl. Meusburger et al 2025).
Die Analysen auf Bestandesebene bestätigten die Ergebnisse der Einzelbaumdaten bis auf den SDI. Obwohl das Einzelbaumwachstum mit zunehmender Bestandesdichte zurückging, nahm der Bestandeszuwachs insgesamt aufgrund der höheren Anzahl Bäume zu. Generell ist bekannt, dass das Waldwachstum zunächst mit zunehmender Bestandesdichte zunimmt, bis ein standortspezifischer Grenzwert erreicht wird, über den das Wachstum aufgrund von Ressourcenlimitierung und Konkurrenz nicht weiter gesteigert werden kann (Pretzsch 2009). In einem gemischten Regressionsmodell mit BFZ des Bestandes als Zielvariablen und dem mittleren BHD, der Bestandesdichte (SDI) sowie der Zeit als interagierende erklärende Variablen ist zu erkennen, dass das Wachstum zwar mit dem SDI zunimmt, dieser Zusammenhang über die Jahre aber abflacht (Abbildung 5). Insgesamt scheint die Wachstumskapazität der Wälder auf den LWF-Standorten in den letzten Jahrzehnten gesunken zu sein, möglicherweise durch abnehmende Wasser- und Nährstoffverfügbarkeit. Dies bestätigen Untersuchungen, die zeigen, dass Wälder mit mittlerer Dichte am besten gegen Trockenheit und Hitze geschützt sind (Sterck et al 2021). Ein abnehmender Trend der Stickstoffdeposition (vgl. Meusburger et al 2025) könnte ebenfalls eine Rolle spielen, wobei noch nicht geklärt ist, ob dies kausal für den abnehmenden Wachstumstrend ist oder ob es sich um eine Koinzidenz handelt.
Zusammenfassung und Ausblick
Die Wachstumsbedingungen unserer heimischen Baumarten ändern sich zunehmend. Die schnellen klimatischen Veränderungen erschweren es langlebigen Organismen wie Bäumen, sich anzupassen. 25 Jahre Waldwachstumsmonitoring zeigen klimabedingte Veränderungen in Wachstum und Bestandesstruktur in den 19 LWF-Wäldern. Besorgniserregend sind die starken negativen Wachstumstrends nicht nur in den Tieflagen, sondern auch in den Hochlagen. Wachstumstrends in dieser Grössenordnung können Auswirkungen auf die forstwirtschaftlichen Erträge, aber auch auf die Schutzfunktion der Wälder haben. Es gilt zu prüfen, ob der Wachstumsrückgang in höheren Lagen ein schweizweites Phänomen ist oder ob die LWF-Wälder aufgrund unterbliebener forstlicher Eingriffe nun an ihre standörtlichen Kapazitätsgrenzen stossen. LFI-Daten der letzten Inventurperiode zeigen zumindest im Alpenraum noch eine Zunahme des Holzvorrats. Allerdings sind auch in Hochlagen bereits negative Auswirkungen und Rückkopplungseffekte durch Klimaveränderungen zu beobachten, nicht nur durch Sturmschäden, sondern auch durch Trockenheit in Boden und Luft (Gharun et al 2020, Trotsiuk et al 2021, Shekhar et al 2024). Der Zusammenhang zwischen Bestandesdichte und Waldwachstum ist auf den LWF-Flächen nach wie vor positiv, allerdings mit abnehmendem Trend, vermutlich durch Ressourcenknappheit, allen voran beim Wasser. So empfehlen Bottero et al (2016) und Andrews et al (2020), die Konkurrenz um Ressourcen durch Ausdünnung des Bestandes zu verringern, um Wälder resistenter gegen die zunehmende Hitze und Trockenheit zu machen.
Wälder sind durch ihre Kohlenstoffaufnahme eine der wichtigsten Kohlenstoffsenken. Dennoch ist zu betonen, dass das Wachstum nur ein Aspekt im Kohlenstoffkreislauf eines Waldbestandes ist (Abbildung 6). So sind im Boden noch grosse Mengen an Kohlenstoff gespeichert, die durch mikrobielle Atmung wieder freigesetzt werden und dessen Dynamiken und Grössenordnungen sich je nach Klimaveränderung stark verändern können (vgl. Hagedorn et al 2025). Ausserdem entziehen Mortalität und Nutzung dem Wald Kohlenstoffvorräte, die hier im Artikel nur am Rande berücksichtigt wurden, die aber gegebenenfalls einen grossen Einfluss auf die Nettokohlenstoffbilanz haben. Die Waldbewirtschaftung steht vor der grossen Herausforderung, die Wälder umfassend anzupassen. Einerseits, um ihre Kapazität als Kohlenstoffsenke weiterhin ausschöpfen zu können, und anderseits, um weitere Ökosystemleistungen wie den Schutz vor Naturgefahren oder die Erholungsfunktion sicherzustellen.

Eingereicht: 15. August 2024, akzeptiert (mit Review): 28. Oktober 2024
Dank
An Christian Hug und das gesamte Sanasilva-Team für die Inventuraufnahmen. An Volodymir Trotsiuk für Datenmanagement und Prozessierung der Wachstumsdaten. An Stephan Zimmermann für die Bereitstellung der chemischen Bodeneigenschaften. An Daniele Pezzotta und das Zentrallabor für die Analysen (Nährstoffe). An Meteotest AG für das Prozessieren und Bereitstellen der Meteodaten und der Stickstoffdeposition. Mit finanzieller Unterstützung des Bundesamtes für Umwelt (BAFU).
Literatur
Low stand density moderates growth declines during hot droughts in semi-arid forests. J Appl Ecol 57 (6): 1089–1102.https://doi.org/10.1111/1365-2664.13615
https://www.bmel.de/DE/themen/wald/wald-in-deutschland/bundeswaldinventur.html. (Zugriff 24.10.2024)
Density-dependent vulnerability of forest ecosystems to drought. J Appl Ecol 54 (6): 1605–1614.https://doi.org/10.1111/1365-2664.12847
Tree mortality in an unmanaged mountain pine(Pinus mugo var. uncinata)stand in the Swiss National Park impacted by root rot fungi. Forest Ecol Manage 145: 79–89.https://doi.org/10.1016/S0378-1127(00)00576-4
Tree growth as indicator of tree vitality and of tree reaction to environmental stress: a review. Europ J Forest Res 124: 319–333.https://doi.org/10.1007/s10342-005-0085-3
European beech responds to climate change with growth decline at lower, and growth increase at higher elevations in the center of its distribution range (SW Germany). Trees 31 (2): 673-686.https://doi.org/10.1007/s00468-016-1499-x
Tree growth in Swiss forests between 1995 and 2010 in relation to climate and stand conditions: recent disturbances matter. Forest Ecol Manage 311: 41–55.https://doi.org/10.1016/j.foreco.2013.05.040
The carbon balance of two contrasting mountain forest ecosystems in Switzerland: Similar annual trends, but seasonal differences. Ecosystems 14: 1289–1309.https://doi.org/10.1007/s10021-011-9481-3
Continental-scale forest growth in Europe is driven by management and further modulated by nitrogen deposition. Forest Ecol Manage 458: 117762.https://doi.org/10.1016/j.foreco.2019.117762
Zurück in die Zukunft – Ein neuer Blick auf die Perspektiven für die Buche nach 20 Jahren Forschung und weiter fortschreitendem Klimawandel. Allg Forst Jagd Z 193: 9–12.https://doi.org/10.23765/afjz000101
Physiological response of Swiss ecosystems to 2018 drought across plant types and elevation. Phil Trans R Soc B: Biol Sci 375 (1810): 20190521.https://doi.org/10.1098/rstb.2019.0521
Kohlenstoff in Schweizer Waldböden – Vorrat und Auswirkung von Trockenheit. Schweiz Z Forstwesen 176 (2): 110–113.https://doi.org/10.3188/szf.2025.0110
Below average midsummer to early autumn precipitation evolved into the main driver of sudden Scots pine vitality decline in the Swiss Rhône valley. Front Ecol Environ 5: 874100.https://doi.org/10.3389/ffgc.2022.874100
Divergent vegetation growth responses to the 2003 heat wave in the Swiss Alps. Geophys Res Lett 32 (8): L18409.https://doi.org/10.1029/2005GL023252
Prognosis and Management Scenarios. In: Brassel P, Lischke H, eds. Swiss National Forest Inventory: Methods and Models of the Second Assessments. WSL Swiss Federal Research Institute for Forest, Snow and Landscape, Birmensdorf. pp197–206.
Klimadaten Waldforschung: Berechnung von historischen Meteo- und Trockenheitsindizes für die Waldforschung auf der Basis von SwissMetNet-Stationsdaten. Bern: Meteotest.
Trends atmosphärischer Deposition und Auswirkungen auf Bodenwasserqualität und Baumernährung. Schweiz Z Forstwesen 176 (2): 92–98.https://doi.org/10.3188/szf.2025.0092
CH2018 - Klimaszenarien für die Schweiz. National Centre for Climate Services, Zürich. pp 24. ISBN 978-3-9525031-0-2
Forest Dynamics, Growth, and Yield. In: Forest Dynamics, Growth and Yield. Springer, Berlin, Heidelberg.https://doi.org/10.1007/978-3-540-88307-4_1
Perfecting a stand-density index for even-aged forests. J Agr Res 46:627– 638.
Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob Change Biol 19: 229–240.https://doi.org/10.1111/gcb.12038
Critical Loads of Nitrogen and their Exceedances. Swiss contribution to the effects-oriented work under the Convention on Long-range Transboundary Air Pollution (UNECE). Federal Office for the Environment, Bern. Environmental studies no. 1642: p 78.
Tree vitality indicators revealed a rapid response of beech forests to the 2018 drought. Ecol Indicators 120: 106903.https://doi.org/10.1016/j.ecolind.2020.106903
Contrasting impact of extreme soil and atmospheric dryness on the functioning of trees and forests. STOTEN 916: 169931.10.1016/j.scitotenv.2024.169931
Optimizing stand density for climate-smart forestry: A way forward towards resilient forests with enhanced carbon storage under extreme climate events. Soil Biol Biochem 162: 108396.https://doi.org/10.1016/j.soilbio.2021.108396
Changements de la végétation herbacée forestière en l’espace de 25 ans sur les sites LWF. Schweiz Z Forstwesen 176 (2): 84–91.https://doi.org/10.3188/szf.2025.0084
Tree growth in Switzerland is increasingly constrained by rising evaporative demand. Global Change Biology 109 (8): 2981–2990.https://doi.org/10.1111/1365-2745.13712